 |
 |
|
|
An effort has been made to strip Behaviour Therapy of the confusion surrounding it , to give it a more solid scientific base. Concepts such as Stimuli and Responses, and Classical and Operant behaviours have been done away with, replaced with the simple notion of a behaviour as being any bodily activity, both external as well as internal; a behaviour being undertaken solely for Positive Reinforcements, either directly or as a consequence of escape from aversive situations.
Learning or Conditioning, it is maintained, is the result of Behaviour and Reinforcement being associated with each other, regardless of the time interval or which comes first, or whether the organism actually carries out the behaviour or actually receives the reinforcement. Discussed also is the perceptual magnification of the reinforcement that takes place after it has first been received.
It is shown that extinction of an undesirable behaviour (whether approach or avoidance) cannot take place unless Counter-Conditioning of the desired behaviour is carried out with a positive reinforcement. Counter-Conditioning thus is the key to behavioural change.
Finally it is emphasized that food or the wrong kind of it, can call forth disturbing thoughts in a client, despite the lack of any positive reinforcement as in masochism, as well as a host of other irrational and self-defeating behaviours.
A behaviour is defined here as any bodily function or activity. This encompasses not only such unconcealed physical movements as walking or lifting an arm but also the acts of looking, listening, smiling, tasting, as well as all those internal goings-on like the beating of the heart or pulse, pressure exerted by the blood, respiration, salivation, sweating, secretions of various glands, etc. etc. etc.
A positive reinforcement (PR) is anything that pleases an organism, and a negative reinforcement (NR) what it displeases.
We now make the following two statements:
(1) THE PURPOSE OF A BEHAVIOUR IS TO GAIN POSITIVE REINFORCEMENT.
(2) FACED WITH A CHOICE BETWEEN TWO OR MORE POSITIVE REINFORCEMENTS, THE ORGANISM TRIES FIRST TO ACQUIRE THE POSITIVE REINFORCEMENT WHOSE VALUE, AS PERCEIVED BY IT, IS GREATER THAN THE VALUES OF OTHER POSITIVE REINFORCEMENTS.
Hungry rats in a T-maze, for example, are seen to prefer that arm of the maze which contains a larger food reward (Sear, 1964).
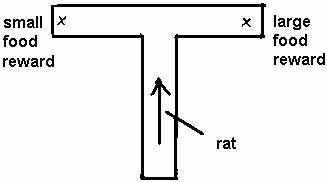
If an electric shock (NR) is then introduced into the preferred arm, its intensity being increased gradually, we would find that at first the animal still prefers the larger reward but that when the shock reaches a certain higher magnitude it switches its preference to the smaller reward (Sear, 1964). That is, the PR derived from escaping the NR (electric shock) is now large enough to combine with the smaller reward to offset the magnitude of the larger reward.
PR (from removal of NR) plus PR (smaller food reward) is greater than PR ( larger food reward).
This 'equation' tells us that if the difference between the two rewards is larger, the intensity of the shock would have to be larger also in order to induce the animal to go to the other arm. That is exactly what Sear (1964) went on to show in his experiment.
We shall define LEARNING or CONDITIONING thus:
LEARNING OR CONDITIONING IS THE OUTCOME OF A BEHAVIOUR THAT IS RELATED TO A REINFORCEMENT.
If we examine the above closely, the following points emerge:
(1) Behaviour (B) and Reinforcement (R) can occur either at the same time or after each other.
(2) The time interval between them (B & R) can be anything.
(3) The organism need not itself perform the behaviour to be learned, nor to receive the reinforcement.
Suppose you were sitting on a bench in a park and you suddenly felt a painful blow struck on your head. If the assailant had come at you from the front, you would have SEEN HIM (B) before, or at the same time as, he hit you (R); this would be a case of FORWARD CONDITIONING and it is usually what happens in real life. If however you had felt the blow from behind, you would have SEEN YOUR ATTACKER (B) after being hit (R), assuming that you had turned your head to look at him; this would be BACKWARD CONDITIONING. In both cases you would have learned to associate a certain person with unpleasantness or to regard him as dangerous. Both forward and backward conditioning are effective so long as the brain keeps being aware of B or R as the case may be or REHEARSES B or R, until R or B happens, respectively. If rehearsal does not take place, for example of B, then R, when it does come, will get attached to a different B -- a behaviour that was not the cause of that R (the genesis of superstition). That is why a reinforcement delivered immediately after a behaviour is more likely to induce learning than one delivered after a period of time. (The R in this example is of course a negative reinforcement, NR. We call the assailant a Conditioned Negative Reinforcement or CNR. The B is the actual behaviour executed by the victim of looking at, or perceiving, the assailant.)
In a study of Backward Conditioning, Keith-Lucas and Guttman (1975) gave rats a foot shock and then presented them with a toy hedgehog -- after time intervals of 1,5,10 and 40 seconds. Only the 40 second group of rats failed to fear the hedgehog, indicating that rehearsal of the shock had ceased in the animals after that long a delay period. In another study by Wagner and Terry (1975) rehearsal was deliberately insured in rabbits by giving them an eye shock when they did not expect it (when something surprising happens to us we keep wondering about it considerably longer than when something usual happens) with the result that backward conditioning was facilitated. Not surprisingly Burkhardt (1980), doing a similar experiment, also showed that larger intensities of shock also produced longer rehearsals (when something very awful happens it lingers more in our memory than when something not so awful happens). Rehearsal, however, can be dispensed with if the organism is made to recollect a bygone B or R. A thief, for example, can be made to reform after punishment delivered long after his crime (punishments are of course usually accompanied by intimations as to why they are meted out). Many of the phobias and other 'neurotic' fears that people suffer from have arisen out of behaviours that were originally harmless but later linked to Negative Reinforcements by society ('My God, did you really do such a disgusting thing?!)
Learning can of course be covertly achieved through any of our senses -- without lifting a finger, so to speak. We can learn about attractive behaviours or unattractive ones simply by listening to what other people tell us or by reading what they have written or by observing what they do (Modelling). We can then go ahead and perform them ourselves (i.e.. if the need for doing so is present in us) or refrain from doing so (to avoid harm).
Since all behaviours are registered in the brain, and all reinforcements too, it is not too much to imagine that it is possible to do a little 'cheating'. We can simply IMAGINE a certain behaviour and then arrange to actually get reinforced for it, or we can carry out a behaviour and then IMAGINE that we had been reinforced for it, or to make it even easier we can IMAGINE both the behaviour and the reinforcement. The result in all three cases would be a strengthening of the bond between B and R, as in ordinary learning. As Cautela and McCullough (1978) explain:
'........the more one thinks of being in an elevator and becoming panicky or being in a plane and a crash occurring, the greater the probability that the individual will engage in avoidance behaviour concerning that particular object. If someone were to imagine that in making a phone call he would stutter when the other person answers, the probability increases that if he makes the call, he will stutter.' (p 241)
Consider deviant sexual behaviour (pedophilia, exhibitionism, voyeurism, etc.). There is evidence (McGuire, Carlisle & Young, 1965) that this comes about through the practice of masturbation (PR) to a memory. Actual experience may have taken place initially (the exhibitionist, for example, achieving a number of orgasms or simply erections while being naked in front of women) but it is the subsequent masturbation to such memories that go to consolidate the deviancy. Here is a man relating how he came to 'appreciate' older women:
'When I was a child of five or six years, we had a live-in housekeeper, as my parents worked in their small department store. The housekeeper must have been forty to fifty years old at the time. My earliest recollection was her putting my penis in her mouth and sucking ever so gently after she bathed me, and I was having a thrill like nothing else. She would always ask me if I felt good, when she knew I finished. By the age of thirteen I was having intercourse as well as oral sex with her. At that time, dad hired more help in the store and mother came home to care for me.
'When I masturbate I fantasize the woman is around sixty years or so (the older the better), and I am a young child and she fondles me and sucks me and I suck her till her body shudders, and she moans like our old housekeeper did.
'To this day, I will only make love to older women (I am forty five and married) and enjoy sex better, much better than with younger women.' (Allan, in Friday, 1980, p29)
In an experiment by Schaefer (1970), monkeys were reinforced with bananas, apples, grapes and peanuts (PR) when they hit themselves on the head (B & NR). At first they were required to bang their heads only a little but gradually the 'demand' was increased until lacerations were produced on the skin. Apart from illustrating the genesis of masochism, this study raises an important question : Since the value of the PR at every delivery did not increase but rather remained the same (as it usually does in such experiments), how come it was still able to offset the very large PR that would have been derived from not punishing oneself so severely?
The reader should put himself in the position of the monkey. If the experimenter told you to bang your head a little against the wall to obtain a certain reinforcement, say a gold watch, you would probably do it; but if he told you to let yourself go and really hurt yourself for the same watch, it is unlikely that you would heed his request. However, having once banged your head a little, it is very probable that you would be prepared to bang it a second time, and a little harder than before, in order to get another watch, and soon, without realizing, you may be hurting yourself increasingly more to obtain the watch again and again, until suddenly you are damaging yourself beyond the limit you were not prepared to exceed initially, despite the reward, the gold watch, not having got any the more valuable.
The fact of the matter is that once a PR has been received, a change occurs in the perception of it. It somehow becomes larger, more attractive than before. It is a psychological phenomenon and there is no other explanation for it. It is tantamount to saying that our need for that PR has increased or that we have been rendered greedy. Perhaps we can now see how an organism is trapped into exposing itself to so much suffering for so little gain (in relation to the original perception).
That there are a mass of behaviours we find ourselves performing without really meaning to is evident from the frequent use of apologetic statements such as 'I am sorry'. We do not give much thought to why a behaviour that is considered reprehensible should have been indulged in in the first place. But the reason simply is REINFORCEMENT. Such is its power.
Having established the mechanism by which learning of all behaviour (appropriate as well as inappropriate) takes place, we shall now look at how we can undo what has been learnt. We call the process COUNTER-CONDITIONING.
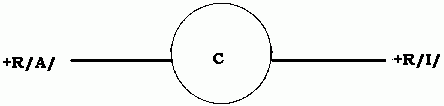
Consider the figure below:
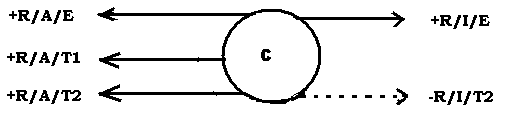
The circle C represents the client who is in need of treatment either because he himself has asked for it or because society demands it. Since all behaviour is caused by the need for a PR and is therefore maintained by the receipt of that PR, we have first +R/I/E, the positive reinforcement that is responsible for the client's inappropriate behaviour (I), the E denoting the fact that it is obtained from his particular environment.
In the behaviourally opposite direction to this is +R/A/E, the positive reinforcement that the client may or may not be aware is ordinarily available from the environment if he behaves appropriately, without the intervention of any therapist, but which, because it is smaller in magnitude (if he is aware of it) than +R/I/E, he does not regard as worth behaving for (recall statement 2).
By behaving inappropriately, he disqualifies himself momentarily from +R/A/E, since the two behaviours are mutually contradictory. For example, if being polite is an appropriate behaviour, not being polite or being rude is an inappropriate behaviour. Of course it can happen that the client is behaving both appropriately and inappropriately at different times and is being reinforced for both behaviours (perhaps in the same way).
The therapist on his own can, on the occurrence of the appropriate behaviour, supply an extrinsic or artificial positive reinforcement, +R/A/T1, and/or on the occurrence of the inappropriate behaviour a negative reinforcement, -R/I/T2.
But -R/I/T2 is an aversion the client would want to avoid by behaving appropriately, and when he does that he is in effect working for a positive reinforcement, +R/A/T2, which is equal in magnitude to -R/I/T2.
We thus have on the diagram all the possible forces affecting the behaviour of a client during therapy. For appropriate behaviour to take place the values of +R/A/E, +R/A/T1 and +R/A/T2 (either on their own or combined with each other) must be greater than the value of +R/I/E.
We might add that in all therapeutic situations, even if the therapist does not tangibly utilize +R/A/T1 and +R/A/T2, these two reinforcements are, to a greater or lesser extent, always present as placebo effects. The former as a consequence of the therapist's helpful presence (Quacks who know nothing about Counter-Conditioning are able to achieve therapeutic results by simply being, in the words of Carl Rogers, 1957, 'warm, empathic and genuine'), the latter as the result of the client's tendency to avoid the therapist's dissatisfaction, perhaps even his censure, by doing what he tells him to do (as for instance in FLOODING --see below-- where the client is made to execute the desirable behaviour in one go, something he had previously been unable to do on his own).
The aim of the behaviour therapist is always to help the client acquire +R/A/E -- if it exists. If not, to prevail upon the environment to make it available to him. The +R/A/T1 that the therapist himself may provide or get the environment to do so for him can only be a temporary solution because of its artificiality in relation to the existing conditions in the environment. However, the -R/I/T2 can be far from temporary since it induces an avoidance behaviour which, if the subsequent environmental contingencies are favourable, can be very persistent indeed, just in the same way that 'naturally' conditioned inappropriate behaviour often is. But this shall be gone into fully under Elimination of Avoidance Behaviour.
A distinction has been made between Approach and Avoidance (inappropriate) behaviours. Approach behaviour is simply any behaviour that satisfies a need of the organism without the organism being in a state of fear. Avoidance behaviour, on the other hand, is anxiety inspired. This, as will be seen, has been done for theoretical reasons. In clinical practice, however, it's not of any importance to classify an inappropriate behaviour into approach or avoidance; the treatment involves the same principle, namely COUNTER-CONDITIONING.
The cessation of a learned behaviour is referred to as EXTINCTION. Here we are concerned with the extinction of non-fearful behaviour.
Imagine, if you will, the following extraordinary situation:
Having committed a serious crime (and society believing as it does in free-will and therefore personal responsibility), you find yourself sentenced to solitary confinement. The cell you are put into has absolutely no windows of any kind through which you could look outside; you are deprived even of the chance to see, or communicate with, your jailers, for your food is delivered quietly through a little opening and your toilet is inside where you are.
The only thing of interest in that little room is a huge television screen sticking out of one of the walls and a small lever beneath it.
You find yourself pulling the lever and lo and behold, the TV screen comes alive. It is as if you were back home in your living room. You settle down to this strange life, consisting of nothing but TV, day in and day out. One day, however, you find that the lever does not work; no amount of pulling or pressing it will light up the screen. Something has gone wrong. Days begin to pass and you feel wretched having to do without the magic screen which was your only stimulation.
The question we want to ask now is this: Will you ever stop pulling that lever?
To be sure, the rate of your lever-pulling will go down with the passage of time as you become more and more 'discouraged', but will it ever cease completely? Remember that you have nothing else to do, except to think, to lie down or to pace the floor. We would therefore expect you once in a while to try the lever 'just to see if it now works'. In other words EXTINCTION of your lever-pulling behaviour would never come about under those special circumstances. And this leads us to the following:
EXTINCTION OF A LEARNED APPROACH BEHAVIOUR CANNOT TAKE PLACE SO LONG AS THE NEED FOR THE PR DERIVED FROM THAT BEHAVIOUR STILL EXISTS, AND NO OTHER BEHAVIOUR WOULD SUPPLY THAT PR.
Thus a hungry rat that has learned to run down a T-maze to obtain food in the right-hand arm, can be made to cease entering that arm only if food is now provided in the left-hand arm. Of course there is such a thing as fatigue, or Reactive Inhibition as Hull (1943) called it, which can stop the animal's search for food even though no alternative behaviour is rewarded but this is not what we mean by Extinction. Such an 'extinction' always produces a SPONTANEOUS RECOVERY of the animal's behaviour at the next experimental session when the fatigue has dissipated. The claims of some writers (e.g.. Lawson, 1960) that Spontaneous Recovery is eliminated with more extinction trials cannot be taken seriously, because with repeated trials the animal simply learns to wait passively until he is taken out of the maze or apparatus to be fed in its home cage, i.e.. it develops an alternative behaviour of remaining still to obtain the same positive reinforcement.
In an experiment by Holz, Azrin, and Ayllon (1963), four chronic psychiatric patients were conditioned to pull a lever for cigarettes. It was found that lever-pulling decreased but did not cease when cigarettes were no longer made available. However, when an alternative response, that of pushing a button, could deliver the cigarettes instead, the subjects abandoned the lever completely.
What is important for true extinction of learned approach behaviour, therefore, is the removal of the derived positive reinforcement and its re-introduction at the execution of an alternative behaviour that is acceptable. (+R/I/E is removed and restored in the form of +R/A/E) Of course +R/A/E may already be present (there are more ways of getting the same thing) in which case only the elimination of +R/I/E is required.
Removal of +R/I/E is not always possible or easy, making it imperative to use the extrinsic +R/A/T1. The client then has a choice between +R/I/E and +R/A/T1, the aim being to eventually phase out +R/A/T1 as +R/A/E, the natural reinforcement for behaving appropriately, takes hold or is appreciated.
Let us consider the experiment by Soloman & Wynne (1953) in which the apparatus, known as a shuttle box, consisted of two compartments separated by a gate. One of the compartments, into which a dog is brought, had a light shining on the ceiling and a grid floor capable of giving an intense electric shock.
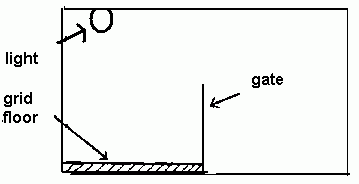
What the experimenters did repeatedly was to turn off the light (CNR) and ten seconds later shock the dog (NR). The animal's initial reaction, as we would expect, was to 'fight back'; it hurled itself around the compartment, smashing into the walls and howling. Very soon it began to display this violent behaviour at the switching off of the light alone (before the shock) as it LEARNED to associate the perception of 'no light' (B) to shock (NR).
Since aggressiveness in such a situation gives no relief (PR), as it does for instance in a victorious combat with another dog, the animal in time switches to what is called a FLIGHT BEHAVIOUR when the light is turned off -- it now wants to run away from the impending shock. In ordinary language we say that the dog has become fearful.
In the shuttle box there is only one possible 'flight' behaviour and that is to jump over the gate to the other compartment where there is no shock. This the animal eventually LEARNS to do and we say that it has undergone a second learning process, namely jumping (B) to avoid shock or to remove its fear (obtain PR).

Suppose now that we were to disconnect the shock apparatus while still presenting the animal with the CNR, thus making the gate-jumping an unnecessary or inappropriate behaviour. How would the dog learn to disregard CNR (or lose its fear) and stay put in the conditioning chamber?
Well, we could try letting it jump again and again until it became so tired (SATIATED) that it could jump no more; or we could physically restrain it from jumping . Our hope in both approaches (which have been given the names EXPERIMENTAL EXTINCTION and RESPONSE PREVENTION, respectively) would be that the dog 'gets wise' to the changed situation and sees no need to continue with the jumping.
Experimental Extinction and Response Prevention do indeed come to make the animal desist from jumping (e.g.. Page & Hall, 1953) and for a time it was believed that this was evidence for the disappearance of fear. But then Page (1955) conducted an experiment in which, after extinguishing the avoidance behaviour of rats in those two ways, he put them in a hungry state in the avoidance compartment, the compartment the animals had run away into during conditioning, and then let them approach food in the conditioning chamber. A third group of rats who had never been exposed to shock was also tested in this way. The eagerness with which these animals approached the shock compartment was expressed in the latencies of 25, 60 and 110 seconds for the control, extinction through satiation and extinction through response prevention, respectively. These figures showed that the experimental animals were still afraid of the shock chamber, and what's more that those who had been restrained retained more fear than those allowed to extinguish through satiation.
It is now known that what really happens is that the organism, having been made to give up an avoidance response, simply acquires other, one might even call them superstitious, responses (such as tics, tremors, struggle, respiratory gasps, crouching, freezing and even grooming) which, as far as the organism is concerned, serve the same purpose as the original avoidance behaviour, namely the removal of fear. A human subject given a traumatic respiratory paralysis might, on entering the area where he had been conditioned (confronted with CNR), for example take 'several deep breaths to re-assure himself' (Campbell, Sanderson, La Verty, 1964, p 638). As Coulter, Riccio and Page (1969) explained:
'Since CS (i.e.. CNR) onset results in a fear response which is reduced by CS offset, then any response S (i.e.. Subject) makes when the CS is terminated will be adventitiously reinforced. As the running response is obstructed during the blocked trials, subjects learn a non-running response (e.g.. crouching, freezing). Immediate or rapid cessation of running on subsequent regular extinction trials would therefore reflect interference from a new response rather than extinction of fear.' (p 380)
It is not always easy to detect such 'superstitious' avoidance responses, in which case an investigation of the organism's internal behavioural state, particularly its heart rate, usually reveals signs of fear (see for example, Werboff, Duane and Cohen, 1964; or Black 1959). It seems that with repeated presentation of the CNR, and therefore with repeated avoidance responses (whatever they may be), the heart rate keeps increasing, We are justified in assuming that this increase reflects an increase in the subjective feeling of fear, also because common experience tells us that the more we avoid a fearful situation the more afraid we seem to become of it (which is why a man who has fallen off a horse is made to mount the animal again quickly in order that he may not risk acquiring a horse phobia). This is in keeping too with what we have said before about perceptual increase in the magnitude of PR -- the PR here being from avoidance behaviour.
If continued avoidance responding, in one way or another, has the effect of increasing fear, we can understand why in the Page (1955) experiment above, the rats that were prevented from escaping were more afraid of the shock chamber than the rats not so restrained. (Latencies of 110 versus 60). A restrained rat has to face the CNR as long as it lasts and thus has to keep repeating its response (for example, trembling) many more times than the single response of running away carried out by the other rats.
In a study by Napalkov (1963) dogs were conditioned after a single pairing of CNR and NR. The CNR was then presented alone repeatedly without the animals being given a chance to escape (a condition analogous to Response Prevention). The following recording of the animals' blood pressure was obtained, plotted against the number of trials (CNR presentations).
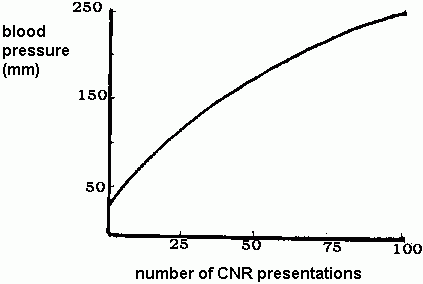
Napalkov reports that this hypertensive state persisted for many months, even after his dogs had been given a rest from the experiment for five months. He realized, however, that this long period of rest was in the presence of stimuli to which the CNR had generalized, making the animals continue to execute their private avoidance responses.
We might add that even if there were no CNRs anywhere in the immediate environment, they could still appear in the imagination through a chain of association with other objects or events. Having established that an organism cannot be made to cease perceiving a CNR as a CNR just by having it remain in its presence without the NR, we are left with the question:
HOW DO WE THEN REMOVE FEAR?
The answer is straightforward:
TURN THE CNR INTO A CPR.
That is to say, we need to present the subject with a Positive Reinforcement while he is being confronted with the CNR, so that the CNR eventually gets COUNTER-CONDITIONED to it (the positive reinforcement). Such an operation has at first the effect of creating in the subject a conflict, a conflict between the need to obtain the positive reinforcement derived from running away from the CNR (fear reduction) and the need to obtain the other positive reinforcement (whatever it may be) that is now being offered to him. As before, we call the former +R/I and the latter +R/A, assuming a therapeutic context.
If we are to succeed, the magnitude of +R/A must be greater than that of +R/I; or we must somehow make sure that the CNR is presented gradually, in stages, so that the value of +R/I remains minimal. The latter approach is usually the more practical. Consider the research work of Wolpe (1958):
A group of cats were individually shocked in a cage just after an auditory stimulus -- a hooter -- was sounded. Since no discrimination training was carried out, the animals were conditioned not only to the hooter but also to the cage, the experimental room as well as to the experimenter himself (this was also true for the experiments of Soloman and Wynne and of Page described above). Their anxiety was manifested, among other things, by their refusal to eat food pellets in the presence of these CNRs and, by Stimulus Generalization, in other rooms of the building.
'Any animal that showed symptoms of tension in room D showed them more strongly in room C, and still more in room B. A cat free of symptoms in room B would also be free in rooms C and D. Thus it would appear that the anxiety-producing effect of these various rooms was a function of their resemblance to room A (the experimental room) as judged in a rough way by the human eye and ear.' (p 53)

The counter-conditioning procedure or the removal of the cats' fears involved finding a sufficiently low intensity of CNR to allow the animals to undertake eating (obtain positive reinforcement) and then to gradually increase this intensity of CNR while maintaining the feeding. Wolpe reports:
'One cat ate initially in room A, one in room B, one in room C, and one in room D. The fifth animal could not be persuaded to eat even in room D but did so eventually in the passage that separated room D from the living cages. Once an animal had eaten in a given place it was given about 20 pellets there, always responding with their presentation with increasing rapidity, and with decreasing signs of anxiety. The next day it was tested in the room next in order of resemblance to room A. By this method of gradual ascent all the animals were eventually enabled to eat in room A. Then, within the room, gradual approach was made to the experimental cage in similar fashion, the animals being fed on the floor increasingly close to the experimental table, then on the table next to the cage, on the roof of the cage, and at last inside the cage. It was found that when feeding became possible in the cage, anxiety reactions were much more rapidly eliminated there when the pellets were tossed at widely distributed points than if they were confined to the food box.' (p 57)
The cats were however still afraid of the hooter for they would not eat when it was sounded.
Counter-conditioning of the hooter was done in two ways: By feeding the animals at decreasing distances to it or by sounding it for increasing periods.
Having now turned all the CNRs into CPRs, the question was: If the PRs (food) suddenly ceased to follow the CPRs, would these CPRs revert back to being CNRs or would they just be neutral stimuli? In other words, would the animals be fearful again?
'The decisive experiment was to extinguish the food-seeking response to the auditory stimulus and then observe whether or not the neurotic reactions were reinstated. Each of the animals was given 30 irregularly massed extinction trials on each of 3 successive days (i.e. hooter followed by no food). Long before the end of the third day's session they all showed almost complete indifference to the auditory stimulus. Immediately after the conclusion of the third extinction session the following test was made. A pellet was dropped on the floor of the experimental cage about two feet away from the animal, and as he began to approach it the auditory signal was sounded continuously, to see if extinction had reinstated the inhibitory effect on eating that had originally been noted. In no instance was there observed any semblance of the restoration of an anxiety response or any suggestion of an inhibition of eating. Moreover, observation for many weeks afterward never revealed occurrence of anxiety responses in any animal.' (p 59)
That was Wolpe with his cats.
Our concern now is to see how such counter-conditioning is applied to human fears. Fortunately the client who comes to the Behaviour Therapist, unlike Wolpe's cats or the dog in the shuttle box, actually WANTS to be able to face the CNR. (Indeed if he didn't he wouldn't present himself for treatment). There is something in the CNR or about it that he feels he must have or tolerate and not being able to do so because he has learnt also to fear it, is the problem. He finds himself in an approach-avoidance situation. For example a person who cannot bring himself to touch other people or shake their hands, because of a fear of dirt or contamination, is depriving himself of normal human intercourse, with all that it entails. +R/I/E, the reinforcement from the inappropriate behaviour of not shaking hands (fear reduction) is greater than +R/A/E, the reinforcement to be derived from having friendly contact with people.
The therapeutic task, therefore, is to alter the balance between +R/A/E and +R/I/E so that the client can undertake the appropriate behaviour (face the CNR) and thus be counter-conditioned. As with cats, if the approach behaviour is carried out gradually, the value of +R/I/E can be kept at zero, allowing the client to experience only 'success' as each step towards his goal is completed. The therapist of course brings his influence to bear in the right direction as when he gives praise and encouragement (+R/A/T1) to the client for undertaking the appropriate behaviour or when he shows disappointment (-R/I/T2) when the client does not make progress. Such verbal reinforcement is often responsible for the reported success of treatment methods other than Behaviour Therapy, without their advocates being aware of it. (See for example Truax, 1966, on Rogerian Psychotherapy)
The above then is the rationale behind the Behaviour Therapy technique of 'Systematic Desensitization'--a name which the reader would now hopefully agree is inaccurate and misleading.
Now if the value of +R/A/T1 were large enough the client could be induced to perform the appropriate behaviour in one fell swoop, so to speak. +R/I/E would then not be able to oppose the combined effects of +R/A/E and +R/A/T1.
To take an example, suppose you were afraid of enclosed places (claustrophobia) and you couldn't bring yourself to go into a movie house (+R/A/E). Suppose then that you fell madly in love with a wonderful person who was crazy about the cinema. The chances are that in order to be with that person (+R/A/T1) you would be prepared to accompany him or her to see a picture, and eventually learn, to your amazement, that your phobia had Gone With The Wind.
One could illustrate such counter-conditioning in animals by getting together fearful and non-fearful ones in the compartment B (fig. below) to which the fearful ones had escaped after being shocked in A, making sure to leave the door between A and B open. Hopefully, the fearful ones would sooner or later be seen mingling with non-fearful ones in the shock compartment A (i.e.. obtaining +R/A/T1). Using then the technique of Page, 1955, we could ascertain the therapeutic results.
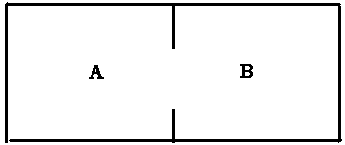
To the best of this writer's knowledge such an experiment has not been done. What has been done (Baum, 1969) is that the fearful ones were 'brutally' dropped into A along with non-fearful ones, with no chance being given to them of escaping to B.
Our equation tells us that this may or may not result in therapeutic gain, depending upon how strong the fear is and how important it is for fearful animals to 'socialize' with non-fearful ones. Baum found signs of counter conditioning, but we can make the statement that if the level of anxiety had been high enough the fearful animals could have been forced to take up 'superstitious' avoidance responses and to forget about socializing, resulting in strengthening of their fear.
But to what extent these superstitious responses would have remained undisturbed by the interaction of other animals is an open question. Perhaps if the experiment were continued for a long enough period, socialization, and therefore counter-conditioning, would have the upper hand.
Whereas the animals in the Baum study had no chance of escape, it is never so with a human being in therapy. A therapist who undertakes -- either imaginally or in vivo -- to expose a client blatantly to a full fledged phobic situation (the procedure has come to be known as FLOODING) always risks being thwarted by the client if the anxiety happens to become unbearable. If the client does not thwart, he may simply 'switch off' his brain, i.e.. refuse to pay attention to the fear evoking situation. Which is why it is not surprising that Hussain, 1971, found that FLOODING seemed to work mostly with people whose anxiety level was not very high. When a client continues with the exposure, while all the time having the power to interrupt it, he is in effect obtaining +R/A/T2 as well (avoiding disappointing the therapist). Note the lack of this for the rats in Baum's experiment.
To take up again our claustrophobia example:
Suppose you happened to inform your loved one about your problem and he or she surprised you by reacting unsympathetically, telling you in effect that you were 'nuts'. This would of course be applying pressure (-R/I/T2) on you, which, if your love was strong enough, you would try to escape from or avoid by stepping into the cinema. Such non-understanding attitudes to odd behaviours may be rare in love relationships but not so in, for example, military service where it is the order of the day. Janet (1925) reports 14 cases of avoidance 'neuroses' improving in the army as a result of the subjects being forced to perform fearful behaviours by regulations.
Baum gave his animals no chance to earn +R/A/T2 but of course it can be arranged. The problem however is that the animals, having first been shocked in the conditioning compartment do not easily learn that the shock has now switched to the safe compartment (so that they may stay put in the conditioning compartment) because repeated avoidance behaviour has magnified their fear of the CNR so much that the shock in the safe compartment (which is invariably of the same intensity as the one used during conditioning) is insufficient to have any effect. Researchers have still not hit upon the idea of employing a higher shock level and, as a result, continue to obtain negative results. What they have managed to find, however, is that by using a different kind of punishment in the safe compartment than that used in the conditioning chamber, it is possible to tilt the balance between the two aversive situations (Kushner & Sandler, 1966) leading to the cessation of avoidance behaviour and to the eventual transformation of the CNR into a CPR.
We might give here a clinical example of FLOODING:
Wolpe (1973):
Mrs. C suffered from agoraphobia (inability to go to open spaces or public places) and it was decided to make her undertake a plane journey alone. She resisted the idea for weeks before giving in (to obtain +R/A/T2). Her husband (+R/A/T1) placed her on the plane and Dr. Wolpe (again +R/A/T1) received her when she alighted from it after the flight (+R/A/E).
Dr. Wolpe reports:
'She had felt increasing anxiety for the first fifteen minutes of the flight, and then gradual subsidence of it. During the second half of the journey she had been perfectly comfortable. She flew home alone the next day without trouble. This single experience resulted in a great increase in her range of comfortable situations away from home.' (p 198-199)
Quite often no instigating CNR is evident from the immediate environment of the clients. The CNR, it seems, occurs inside their brains, through the mechanism of association (of one thought with another). Such an imaginal appearance of the CNR ordinarily poses no problem for anybody: what we do is to simply brush it aside and think of something else. But some people are unable to do this and as a consequence are forced to be anxious and to attempt a variety of behaviours, some even bizarre, to free themselves from it -- all to no avail. The phenomenon is referred to as Intrusive or Obsessive thoughts and it at first sight goes counter to what we have said about organisms being attracted to positive reinforcements and repelled by negative reinforcements, for why should a person persist in entertaining thoughts that make him so unhappy? One could understand if these thoughts were followed by positive reinforcements (the mechanism of masochism) but it is not the case here. The most logical explanation is that the brains of these people are being STIMULATED, letting their past CNRs invade into the consciousness and remain there as well as making them more sensitive to subsequent NRs. There is in fact a good deal of evidence for this.
Stimulation can be effected in three main ways:
Electrical, Chemical and Nutritional.
And besides being in the negative direction, it can also be in the positive direction, i.e.. the CPRs too can be brought to the fore in the psyche.
In their pioneering work, Olds and Milner (1954) planted electrodes in the septal area of the forebrains of rats so that they could be used to deliver electric current after the animals had recovered from surgery. It was shown that such stimulation acted as a PR since, having been delivered only when the rat came into a particular area of the cage, it made the animal's entry into that area increasingly more frequent. If a similar electrical stimulation were carried out on a human being what we would expect is a show of gaiety on the part of the subject. This is indeed what Heath (1964) has reported:
'Subjects were not informed when stimuli were applied. When questioned concerning changes in mental content, they were generally at a loss to explain them. For example, one patient on the verge of tears described his father's near-fatal illness and condemned himself as somehow responsible, but when the septal region was stimulated, he immediately terminated this conversation and within 15 seconds exhibited a broad grin as he discussed plans to date and seduce a girl friend. When asked why he had changed the conversation so abruptly, he replied that the plans concerning the girl suddenly came to him. This phenomenon was repeated several times in the patient: stimulation was administered to the septal region when he was describing a depressive state, and almost instantly he became gay. Another severely agitated and depressed subject whose verbalizations expressed self-condemnation and hopelessness (a condition that had prevailed for over 2 years) smiled broadly and related a sexual experience of his youth within one minute after onset of septal stimulation.' (p 224)
That there are other areas of the brain the stimulation of which acts like a punishment has been shown by Delgado, Roberts, and Miller (1954). They happened to work with cats.
It is of course well known that chemicals influence the mind; some, like the so-called tranquilizing drugs, in a positive way; others, such as lactic acid (Pitts & McClure, 1967), in a negative way.
It is known also that listening to music, looking at certain colours, being rocked in a hammock or a chair and physical exercise promote a feeling of calmness and well-being.
But what is more important is that the food we eat has a lot to say about what we think and do. Conditions such as depression, hypochondria, floating anxieties, schizophrenia, lack of empathy, OCD, etc (all reflecting the kind of CNRs one has acquired) are more likely to be the result of powerful inner instigators, with food (or the wrong kind of it) as the principal culprit, and therefore beyond easy reach of purely external reinforcements.
There have been several studies on the role of food in mental disturbance. What we can do here is to reproduce an abstract of a paper by Gettis, 1989, which is a review of the various studies. Gettis writes:
"The literature dealing with the relationship between food sensitivities and psychological disturbance is reviewed. Numerous theorists and researchers believe the problems of persons presenting with adverse reactions to foods are psychological when immunological techniques fail to confirm an allergic basis. However, there is mounting evidence that adverse reactions to foods can most likely be caused by a variety of mechanisms, and that food sensitivities may indeed cause or exacerbate symptoms of a psychological nature."
One wonders what it is that food allergy really does to induce mental disturbance? A limited study by Rippere, 1984, seems to suggest that obsessive ruminations are probably linked to hypoglycaemia; and we know that the brain cannot function properly without an adequate amount of sugar being supplied to it; so it is reasonable to suspect that food allergy may be having the effect of depriving the brain of sugar or of causing hypoglycaemia. There is further backing to this suspicion from research on physical exercise: it has been shown (see link below) that the brain receives an increased flow of sugar as a result of exercise. Since physical exercise is of course well recognized as being beneficial to psychiatric patients (see link below ), we are perhaps justified in supposing that sugar or the deficiency of it, may be the determining factor in mental disturbance. Writers like Carlton Fredericks (Psycho-Nutrition, Jan 83) have long claimed that food allergy can cause hypoglycaemia and that hypoglycaemia can in turn cause food allergies. The subject needs scientific investigation.
Behaviour Therapists, it must be said in conclusion, need to investigate the client's eating and drinking habits and order a change of diet and daily physical exercise such as jogging before carrying out the more permanent solution of Counter-Conditioning.
Baum, M. 'Extinction of an avoidance response motivated by intense fear: social facilitation of the action of response prevention (flooding) in rats.' Behaviour Research and Therapy. 7,57-62,1969.
Black, A.H. 'Heart rate changes during avoidance learning in dogs.' Canadian J. Psychology. 13,229-242, 1959.
Burkhardt, P.E. 'One-trial backward fear conditioning in rats as a function of US intensity.' Bulletin of the Psychonomic Society. 15,9-11,1980.
Campbell, D; Sanderson, R.E.; Laverty, S.G. 'Characteristic of a conditioned response in human subjects during extinction trials following a simple traumatic conditioning trial.' J. Abnormal Soc. Psychology. 68,627-639, 1964.
Cautela, J.R.; McCullough, L. 'Covert Conditioning' in THE POWER OF HUMAN IMAGINATION (Ed) Singer, J.L.; Pope, K.S. Plenum Press, NY, 1978.
Coulter, X; Riccio, D.C.; Page, H.A. 'Effects of blocking an instrumental avoidance response.' J. Comp. & Physo. Psychol. 68,3,377-381,1969.
Delgado, J.M.R.; Roberts, W.W.; Miller, N.E. 'Learning motivated by electrical stimulation of the brain.' Amer. J. Physiology. 179,587-593,1954.
Friday, N. Men in Love. Hutchinson, London, 1980.
Gettis,A. Food sensitivities and psychological disturbance: a review. Nutr. Health 6(3): 135-46, 1989.
Heath,R.G. 'Pleasure response in human subjects to direct stimulation of the brain: Physiologic and Psychodynamic considerations.' In R.G. Heath (Ed) The role of pleasure in behaviour. NY. Hoeber, 1964.
Holz, W.C.; Azrin, N.H.; Ayllon, T. 'Elimination of behaviour of mental patients by response produced extinction.' J. Exp. Analysis of Behaviour. 6,3,407-412,1963.
Hull, C.L. Principles of Behaviour. NY. Appleton Century Crofts. 1943
Hussain, M.Z. 'Desensitization and flooding (implosion) in treatment of phobias' Amer. J. Psychiatry. 127,11,May 1971.
Janet, P. Psychological Healing. NY. Macmillan. Vol.2,1925.
Keith-Lucas, T; Guttman, N. 'Robust single-trial delayed backward conditioning' J. Comp. & Physiolo. Psycholo. 88,468-476,1975.
Kushner, M.; Sandler, J. 'Aversion therapy and the concept of punishment' Beh. Res. & Therapy. 4,179-186,1966.
Lawson, R. Learning and Behaviour. NY. Macmillan. 1960.
McGuire, R.J.; Carlisle, J.M.; Young, B.C. 'Sexual deviations as conditioned behaviour: a hypothesis.' Beh. Res. & Therapy. 2,185-190,1965.
Napalkov, A.J. 'Information processing of the brain.' In Progress of Brain Research. (Ed) Wiener, N. & Sefade, J.C. Vol. 2,59-69,1963.
Olds, J.; Milner, P. 'Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain.' J.Comp. Physiolo. Psycholo. 47,419- 427,1954.
Page, H.A. 'The facilitation of experimental extinction by response prevention as a function of the acquisition of a new response.' J. Comp. Physiolo. Psycholo. 48,14-16,1955.
Page, H.A.; Hall, J.F. 'Experimental Extinction as a function of the prevention of a response.' J. Comp. Physiolo. Psycholo. 46,33 34,1953.
Pitts, F.N.; McClure, J.N. 'Lactate Metabolism in Anxiety Neurosis.' New England J. Med. 277,1329-1336,1967.
Rippere,V. "Can hypoglycaemia cause obsessions and ruminations?" Med Hypothesis. 1984 Sept; 15(1); 3-13.
Rogers, C.J. Consulting Psychology. 21,95- ,1957.
Schaefer, H.H. 'Self-injurious behaviour: shaping head banging in monkeys.' J. Applied Behaviour Analysis. 3,111 116,1970.
Sears, D.O. 'Punishment and choice in the rat.' J. Comp. Physiolo. Psycholo. 57,297-299,1964.
Soloman, R.L., Wynne, L.C. 'Traumatic avoidance learning: acquisition in normal dogs.' Psychological Monographs. 67,4,1-19,1953.
Truax, C.B. 'Reinforcement and non-reinforcement in Rogerian Psychotherapy.' J. Abnormal Psychology. 71,1-9,1966.
Wagner, A.R.; Terry, W.S. 'Backward conditioning to a CS following an expected vs. a surprising UCS.' Animal Learning and Behaviour. 3,370-374,1975.
Werboff, J.; Daune, D.; Cohen, B.D. 'Extinction of conditioned avoidance and heart rate responses in rats.' J. Psychosomatic Research. 8,29-33,1964.
Wolpe, J. Psychotherapy by Reciprocal Inhibition. Stanford University Press. Stanford. California. 1958.
Wolpe, J. The practice of behaviour therapy. NY. Pergamon Press. 1973.
Also by the author: